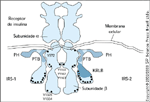
  FIGURA 2-3 FIGURA 2-3
Modelo para a interaçăo dos receptores de insulina (IR – insulin receptors) e moléculas de substrato do receptor de insulina (IRS – insulin receptor substrate). As proteínas IRS foram identificadas como proteínas fosforiladas em tirosina nas células tratadas com insulina [26–28]. Além da insulina, outros agentes como os fatores de crescimento semelhantes ŕ insulina e interleucinas estimulam a fosforilaçăo de proteína IRS [20]. Após ativaçăo da IR quinase, o domínio de ligaçăo ŕ fosfotirosina (PTB – phosphotyrosine-binding) do IRS torna-se intimamente associado ŕ regiăo justamembranar do IR. Esta interaçăo exige fosforilaçăo da tirosina 972 no IR. Além disto, é possível que o domínio de homologia da plecstrina (PH) estabilize esta interaçăo. O domínio PH está localizado na extremidade aminoterminal da proteína IRS, que poderia promover a interaçăo com a bicamada membranar. Muitos domínios PH se ligam a fosfolípides. Além disto, existe pelo menos um domínio adicional que interage com o IR, chamado domínio de ligaçăo ŕ alça regulatória da quinase (KRLB – kinase regulatory loop-binding). Sua interaçăo com o IR exige fosforilaçăo de resíduos de tirosina no domínio catalítico do receptor (tirosinas 1158, 1162 e 1163). Por estar presente apenas em Irs-2, o KRLB poderia determinar especificidade da interaçăo entre o IR e seus substratos [29,30]. Experimentos de ablaçăo gęnica em camundongos indicam que as várias proteínas IRS tęm funçőes singulares. Camundongos nos quais Irs-1 está ausente apresentam retardo de crescimento e săo discretamente resistentes ŕ insulina, sugerindo que o Irs-1 desempenha um importante papel na funçăo do receptor tanto da insulina quanto do fator-1 de crescimento semelhante ŕ insulina; camundongos com ausęncia de Irs-2 desenvolvem resistęncia ŕ insulina e apresentam desenvolvimento comprometido de células b, sugerindo que o Irs-2 é importante para regulaçăo metabólica em resposta ŕ insulina [4,21].
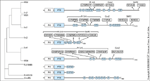 FIGURA 2-4 FIGURA 2-4
Estrutura das proteínas IRS. As proteínas IRS năo tęm atividades catalíticas intrínsecas, mas săo compostas de vários domínios de interaçăo e de motifs de fosforilaçăo. Ocorrem pelo menos tręs proteínas IRS em camundongos e pessoas, que incluem o IRS1 e o IRS2 que săo amplamente expressos, enquanto o IRS4 está limitado ao timo, cérebro, rim e, possivelmente, células b pancreáticas [31]. Os roedores também expressam o Irs3, que está basicamente restrito ao tecido adiposo e exige uma atividade semelhante ao do Irs1; entretanto, é possível que este curto ortolog năo ocorra em seres humanos [32]. A análise filogenética revela uma íntima relaçăo evolutiva entre IRS1/Irs1 e IRS2/Irs2 de pessoas/camundongos, que podem ter divergido a partir de IRS4/Irs4. A proteína IRS de Drosophila, Chico, está fracamente relacionada com seus ortologs mamíferos já que contém poucos sítios de fosforilaçăo de tirosina terminais. Finalmente, a análise de seqüęncia do genoma humano revela pelo menos 2 proteínas IRS putativas reconhecidas por domínios adjacentes de homologia da plecstrina (PH) e de ligaçăo ŕ fosfotirosina (PTB); entretanto, elas contęm caudas COOH bem curtas com poucos sítios de fosforilaçăo da tirosina e, portanto, sua funçăo permanece desconhecida (veja a Fig. 2-2).Todas as proteínas IRS săo caracterizadas pela presença de um domínio PH NH2-terminal adjacente a um domínio PTB, seguidos por uma cauda COOH-terminal de comprimento variável que contém inúmeros sítios de fosforilaçăo de tirosina e de serina. Os domínios PH e PTB medeiam interaçőes específicas com as quinases de receptor da insulina e do fator-1 de crescimento semelhante ŕ insulina (IGF-1) [33,34]. O domínio PTB se liga a motifs NPXY fosforilados nos receptores da insulina, do IGF-1 ou da interleucina-4; entretanto, outros receptores que promovem a fosforilaçăo de tirosina da proteína IRS năo contęm motifs NPXY [35]. Os candidatos a parceiros de ligaçăo ao domínio PH poderiam incluir fosfolípides, peptídios ácidos ou proteínas específicas como PHIP [36,37]. Entre as proteínas que comprovadamente se ligam ao IRS1, a mais proeminente é a subunidade p85 da fosfatidilinositol 3-quinase (PI 3-K), cuja atividade enzimática acredita-se que seja crucial na estimulaçăo de muitas açőes biológicas da insulina.
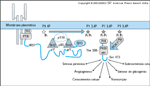 FIGURA 2-5 FIGURA 2-5
Ocorre fosforilaçăo de tirosina em vários motifs de seqüęncia de aminoácidos que criam sítios de ligaçăo para os assim chamados motifs SH2 (de src homology 2) [38]. Os motifs SH2 tęm 50 a 100 aminoácidos de comprimento e se ligam com alta afinidade a resíduos de fosfotirosina em várias moléculas de sinalizaçăo intracelular. Durante a estimulaçăo insulínica, as proteínas de substrato do receptor da insulina (IRS – insulin receptor substrate) fosforiladas em tirosina se ligam aos dois domínios SH2 na subunidade regulatória (p85) da fosfatidilinositol 3-quinase (PI 3-K), resultando na ativaçăo da subunidade catalítica p110. A subunidade p110 catalisa a fosforilaçăo do fosfatidilinositol (PI) na posiçăo D3 do anel de inositol, levando ŕ geraçăo de PI 3-fosfato a partir de PI, de PI 3,4-bisfosfato a partir de PI 4-fosfato e de PI 3,4,5-trisfosfato a partir de PI 4,5-bisfosfato. Acumulam-se as evidęncias de que os inositídeos fosforilados em D3 agem como mensageiros intracelulares que levam ŕ ativaçăo de quinases dependentes de PI, alteraçőes no tráfego intracelular e estimulaçăo do crescimento [39]. Os produtos da PI 3-quinase interagem com serina quinases citosólicas (PDK, PKB e outras) para gerar um sistema de sinalizaçăo ligada ŕ membrana que medeia diversos processos biológicos, inclusive síntese de glicogęnio e transporte de glicose. O domínio PH da PKB liga-se ao PI 3-fosfato, mas uma atividade enzimática máxima exige fosforilaçăo da Thr308 e da Ser473 mediada pela PDK [40]. Além de seu papel na regulaçăo metabólica, a PKB promove a inibiçăo da apoptose mediada pelo fator-I de crescimento semelhante ŕ insulina, bem como a ativaçăo da p70S6k quinase com as concomitantes síntese proteica, regulaçăo da entrada no ciclo celular e diferenciaçăo celular [41]. PKB — proteína quinase B; PDK — proteína quinase dependente de fosfoinositídeo.
voltar |
















;)
;)
;)