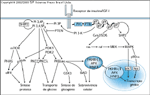
 FIGURA 2-6 FIGURA 2-6
As proteínas de substrato do receptor da insulina (IRS – insulin receptor substrate) coordenam várias vias de sinalizaçăo a jusante. A diversidade da açăo da insulina em diferentes tecidos é parcialmente explicada pelos diferentes produtos acoplados ŕs vias ativadas pelo hormônio. Há dois ramos principais que propagam o sinal gerado através do receptor da insulina: o ramo da fosfatidil 3-quinase (PI 3-K) e o ramo da proteína quinase ativada por Ras/mitógeno. As proteínas IRS se ligam a PI 3-K, Grb2/SOS e SHP-2. O complexo GRb2/SOS medeia a ativaçăo de p21ras, ativando desta forma a cascata ras raf metiletilcetona (MEK) proteína quinase ativada por mitógeno (MAP). O SHP2 retroalimenta para inibir a fosforilaçăo de proteína IRS através da desfosforilaçăo direta da proteína IRS, mas também poderia transmitir um sinal independente para ativar a MAP quinase. A MAP quinase ativada fosforila p90rsk que, por sua vez, fosforila o fator de transcriçăo c-fos, aumentando sua atividade transcricional. Da mesma forma, a MAP quinase fosforila elk1, aumentando sua atividade transcricional. A ativaçăo da PI 3-K pelo recrutamento da proteína IRS resulta na geraçăo de PI 3,4P2 e PI 3,4,5P3 (antagonizada pela açăo de PTEN ou SHIP2). Juntos, PI 3,4P2 e PI 3,4,5P3 ativam várias quinases, inclusive mTOR, isoformas atípicas de PKC e proteína quinase dependente de fosfoinositídeo (PDK1). A PDK1 está a montante de PKB, o que promove o transporte de glicose; as isoformas atípicas de PKC também desempenham um papel. A PKB também regula GSK3, que poderia regular a síntese de glicogęnio e uma variedade de reguladores de sobrevida celular; a fosforilaçăo de BAD mediada pela PKB inibe a apoptose. A PKB também fosforila os fatores de transcriçăo 'forkhead' (como o Foxo1), que restringe sua localizaçăo ao citoplasma e inibe sua atividade transcricional. PKB — proteína quinase B; GAP — proteína associada ŕ guanosina trifosfatase; GLUT4 — transportador-4 de glicose; GRB-2 — proteína-2 de ligaçăo ao receptor do fator de crescimento 2; GSK3 — glicogęnio-sintase quinase 3; MAPKK — MAP quinase quinase; PKC — proteína quinase C; SOS — son-of-sevenless. raf metiletilcetona (MEK) proteína quinase ativada por mitógeno (MAP). O SHP2 retroalimenta para inibir a fosforilaçăo de proteína IRS através da desfosforilaçăo direta da proteína IRS, mas também poderia transmitir um sinal independente para ativar a MAP quinase. A MAP quinase ativada fosforila p90rsk que, por sua vez, fosforila o fator de transcriçăo c-fos, aumentando sua atividade transcricional. Da mesma forma, a MAP quinase fosforila elk1, aumentando sua atividade transcricional. A ativaçăo da PI 3-K pelo recrutamento da proteína IRS resulta na geraçăo de PI 3,4P2 e PI 3,4,5P3 (antagonizada pela açăo de PTEN ou SHIP2). Juntos, PI 3,4P2 e PI 3,4,5P3 ativam várias quinases, inclusive mTOR, isoformas atípicas de PKC e proteína quinase dependente de fosfoinositídeo (PDK1). A PDK1 está a montante de PKB, o que promove o transporte de glicose; as isoformas atípicas de PKC também desempenham um papel. A PKB também regula GSK3, que poderia regular a síntese de glicogęnio e uma variedade de reguladores de sobrevida celular; a fosforilaçăo de BAD mediada pela PKB inibe a apoptose. A PKB também fosforila os fatores de transcriçăo 'forkhead' (como o Foxo1), que restringe sua localizaçăo ao citoplasma e inibe sua atividade transcricional. PKB — proteína quinase B; GAP — proteína associada ŕ guanosina trifosfatase; GLUT4 — transportador-4 de glicose; GRB-2 — proteína-2 de ligaçăo ao receptor do fator de crescimento 2; GSK3 — glicogęnio-sintase quinase 3; MAPKK — MAP quinase quinase; PKC — proteína quinase C; SOS — son-of-sevenless.
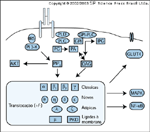 FIGURA 2-7 FIGURA 2-7
Membros da família PKC das serina/treonina quinases estăo implicados em várias açőes da insulina. Há dois subgrupos de PKCs. As isoformas PKC “clássicas” se ligam ao cálcio para ativaçăo. As PKCs atípicas săo ativadas pela ligaçăo com o diacilglicerol (DAG) ou com outros fosfolípides, como fosfatidilinositol 3,4,5-trisfosfato (PIP3). A insulina ativa diferentes membros desta família de quinases através da formaçăo de DAG e PIP3. A insulina estimula a formaçăo de DAG através da hidrólise da fosfatidilcolina (PC) em DAG e ácido fosfatídico (PA) ou através da atividade de uma fosfolipase-C específico de glicosil-fosfatidilinositol (GPI-PLC), levando ŕ formaçăo de DAG inositol-fosfoglicana (IPG). Foi demonstrado que diferentes formas de PKC sofrem translocaçăo do citosol para a membrana em resposta ŕ estimulaçăo da insulina em diferentes tecidos. Existem evidęncias de que este processo possa ser importante para a atividade biológica das PKCs [42]. Sabe-se que as PKCs podem ativar diretamente a via da MAP quinase e o fator de transcriçăo NF-kB, levando a um aumento de expressăo gęnica e síntese proteica. Mais recentemente, emergiram evidęncias de que a ativaçăo de PKCs atípicas pelo PIP3 possa ser importante no processo de transporte de glicose insulino-dependente [43].
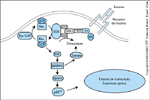 FIGURA 2-8 FIGURA 2-8
A via de sinalizaçăo Ras desempenha um papel importante no crescimento e transformaçăo celular [38]. A insulina ativa a via Ras, levando a uma maior síntese proteica e mitogęnese. A ativaçăo do Ras em resposta ŕ insulina exige a formaçăo de um complexo de sinalizaçăo entre o substrato do receptor da insulina (IRS – insulin receptor substrate) ou shc (src homology 2/collagen homology-containing protein) e a proteína-2 de ligaçăo ao receptor do fator de crescimento (GRB-2 – growth factor receptor binding protein 2). A GRB-2 consiste em um domínio SH2 que se liga aos resíduos fosforilados de tirosina no IRS ou shc, e em dois domínios SH3 (src homology 3 motif) que reconhecem motifs ricos em prolina no fator de ‘permuta son-of-sevenless’ (SOS). O SOS cataliza a conversăo do Ras de uma forma inativa (ligada ao difosfato de guanosina) para uma forma ativa (ligada ao trifosfato de guanosina). Uma rasGAP catalisa a reaçăo reversa, com a formaçăo de Ras inativo. Ras-GTP ativa a Raf quinase, que fosforila a quinase da proteína quinase ativada por mitógeno (MAPKK). A MAPKK fosforila quinases de regulaçăo extracelular Erk1/2, que estimulam outras quinases, inclusive p90rsk, que promove síntese proteica e expressăo gęnica. Ao ser ativada, a MAPKK também ativa vias adicionais que catalisam a dissociaçăo de GRB-2 do SOS, que finaliza o sinal. Parece que os papéis relativos do IRS e shc na ativaçăo de SOS através de GRB-2 variam em diferentes tecidos [44].
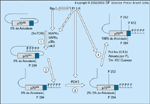 FIGURA 2-9 FIGURA 2-9
A insulina regula a p70s6 quinase, que promove síntese proteica e proliferaçăo celular. A p70s6k ocorre como duas isoformas denominadas S6K1 e S6K2. Um distúrbio em s6k1 causa intolerância ŕ glicose por causa do tamanho reduzido das células b de ilhota; em contraste, a açăo periférica da insulina poderia ser amplificada, sugerindo que a S6K1 inibe a sinalizaçăo da insulina [45]. A S6K1 será ativada in vivo através de eventos de fosforilaçăo em múltiplos sítios em resposta ŕ insulina e a outros mitógenos se estiver disponível um suprimento suficiente de aminoácidos ou em resposta a altas concentraçőes de aminoácidos isoladamente [46]. Portanto, acredita-se que a p70s6k seja um sensor de nutriente [47]. Uma variedade de quinases direcionadas pela prolina, entre elas a Erk, a Cdc2 e a mTOR, fosforila vários sítios na p70s6k, inclusive sítios Ser/Thr-Pro no domínio auto-inibitório de pseudo-substrato da cauda carboxiterminal (Ser434, Ser441, Thr444, Ser447 e Ser452) bem como sítios na extensăo do domínio catalítico (Thr390 e Ser394) [46]. A fosforilaçăo do domínio auto-inibitório de pseudo-substrato serve para desengatar a cauda carboxiterminal, possibilitando o acesso a Thr252 e Thr412. A PDK1 catalisa a fosforilaçăo de Thr252 na alça de ativaçăo do domínio catalítico, que aumenta ligeiramente a atividade da quinase e facilita a fosforilaçăo de Thr412 estimulada por PtdIns (3,4,5)P3, que é a etapa limitante de velocidade na ativaçăo. A fosforilaçăo concomitante de Thr252 e Thr412 aumenta a atividade da p70s6k em 20 a 30 vezes [39]. A desativaçăo da p70s6k que ocorre com a retirada da insulina ou a inibiçăo da PI 3 quinase resulta da desfosforilaçăo total de Thr412, com fosforilaçăo residual remanescente em Thr252 refletindo a necessidade de (412/252) fosforilaçăo dual para a atividade completa da quinase.
voltar |
















;)
;)
;)
;)