  FIGURA 15-6 FIGURA 15-6
Requisito para um peso estável. Um peso estável exige uma equivalęncia entre calorias ingeridas e calorias gastas. A massa livre de gordura é um importante determinante de gasto de energia em repouso [6]. O ganho de peso leva a um aumento tanto na massa gordurosa quanto na massa livre de gordura; portanto, a taxa metabólica basal de pessoas obesas é maior que a de pessoas magras da mesma altura [7]. A obesidade resulta do excesso crônico de calorias ingeridas em relaçăo ŕs calorias gastas. As calorias gastas incluem a taxa metabólica em repouso, os efeitos térmicos do alimento e exercícios e a termogęnese adaptativa. A gordura corpórea năo é significativamente influenciada pelo gasto de energia em repouso nem pelo efeito térmico do alimento [8], mas alteraçőes no gasto de energia resultante de atividade física influenciam o peso e a composiçăo corpórea [9].
 FIGURA 15-7 FIGURA 15-7
Homeostase de energia. Durante muitos anos, a obesidade foi considerada a conseqüęncia de um defeito moral. Comer era considerado um processo inteiramente sob o controle voluntário e gasto reduzido de energia era atribuído ŕ inatividade. Com o tempo, diversos estudos revelaram que a termogęnese variava entre as pessoas e que, quando colocada em dietas compostas de calorias iguais, as pessoas poderiam ganhar, manter ou até perder peso. A descoberta da leptina em 1994 revolucionou a compreensăo da base fisiopatológica da obesidade. É agora evidente que vários fatores regulam o apetite e o gasto total de energia. A demonstraçăo de que defeitos gęnicos únicos podem levar ŕ obesidade também demonstrou que a alimentaçăo excessiva é um processo que nem sempre é acessível ao controle consciente.
 FIGURA 15-8 FIGURA 15-8
Organizaçăo hipotalâmica. O papel do hipotálamo na regulaçăo do comportamento alimentar foi inicialmente definido há décadas em estudos de estimulaçăo elétrica e lesőes anatômicas. O papel do hipotálamo lateral na mediaçăo da alimentaçăo foi considerado pela primeira vez em 1951 [10]. Em 1940, foi relatado que lesőes hipotalâmicas experimentais do hipotálamo ventromedial produziam obesidade [11].
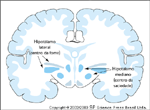 FIGURA 15-9 FIGURA 15-9
Áreas hipotalâmicas implicadas no comportamento alimentar. O hipotálamo lateral é verde (a área em verde mais claro indica a zona incerta). O hormônio concentrador de melanina [12] e a orexina [13] situam-se nesta área e estimulam a fome. O hipotálamo mediano (ventromedial e dorso-medial) é vermelho e o núcleo paraventricular é amarelo. O núcleo arqueado, onde estăo localizados os corpos celulares que produzem o neuropeptídio Y, a preproopiomelanocortina (o precursor do hormônio que estimula melanócitos), o peptídio relacionado a agouti e o transcrito regulado por cocaína e anfetamina [CART – cocaine- and amphetamine-regulated transcript], é rosa. O neuropeptídio Y e o peptídio relacionado ao agouti estimulam a ingestăo alimentar e o hormônio estimulador de melanócitos e o CART inibem a ingestăo alimentar.
As monoaminas também desempenham um papel na regulaçăo do apetite. Em animais experimentais, a administraçăo de noradrenalina leva a um aumento agudo na ingestăo de alimento e a administraçăo crônica leva ao ganho de peso. Este efeito parece ser dependente do receptor estimulado porque os agonistas a2 aumentam a ingestăo de alimento enquanto os agonistas a1 e b2 levam a uma reduçăo na ingestăo alimentar [14-16]. A dopamina desempenha um papel importante na regulaçăo da ingestăo alimentar; entretanto, os efeitos dependem do lugar em que ela é liberada. A administraçăo de dopamina no hipotálamo lateral leva a uma refeiçăo de menor tamanho [17] enquanto a liberaçăo no núcleo accumbens aumenta a ingestăo de alimento [18]. A serotonina também afeta a ingestăo de alimentos; entretanto, os efeitos săo complexos e variam dependendo de qual área anatômica do cérebro é o alvo e do receptor ativado. 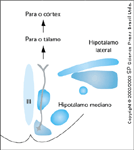 FIGURA 15-10 FIGURA 15-10
Neuropeptídio Y. O neuropeptídio Y (NPY) é produzido nos neurônios do núcleo arqueado. O papel do NPY na alimentaçăo é atribuído a projeçőes diretas do NPY ao núcleo paraventricular [19]. Injeçőes repetitivas de NPY no hipotálamo induz hiperfagia e obesidade [20]. O neuropeptídio Y também altera o metabolismo de energia. Depois de repetidas injeçőes de NPY, a atividade termogęnica da gordura castanha está diminuída e a atividade da lipoproteína lipase da gordura branca está diminuída [21]. Este achado sugere que os neuropeptídios que regulam o apetite podem ter múltiplos papéis.
Uma das açőes da leptina é a supressăo da síntese de NPY no núcleo arqueado. Este peptídio parece funcionar como um importante regulador central do comportamento alimentar. A injeçăo de NPY nos ventrículos laterais de rato leva a um acentuado aumento na alimentaçăo; animais tratados com NPY comem seis a dez vezes mais que animais controle durante o período de 24 horas seguintes. Injeçőes repetidas durante vários dias causam ganho de peso. O neuropeptídio Y também suprime o gasto de energia através de açőes sobre o sistema nervoso simpático. Embora o NPY seja difusamente expresso, săo os neurônios sintetizadores de NPY do núcleo arqueado que se projetam para o núcleo paraventricular e o hipotálamo dorsomediano que săo responsáveis pela mediaçăo do comportamento alimentar. No camundongo ob/ob, os níveis de RNA mensageiro (mRNA) no núcleo arqueado săo duas a tręs vezes maiores que os níveis de mRNA nos camundongos controle; os níveis do peptídio estăo também aumentados no camundongo ob/ob. Resultados semelhantes săo encontrados no rato Zucker gordos. Tanto o tratamento intracerebroventricular central quanto o periférico com leptina de animais ob/ob reduziram as mensagens de NPY no núcleo arqueado, sugerindo que o NPY seja um alvo da leptina. Em animais normais, a administraçăo periférica de leptina inibe significativamente o aumento nos níveis de mRNA de NPY do arqueado observados com a inaniçăo. Embora o NPY possa normalmente mediar o comportamento alimentar, camundongos com knockout de NPY nos quais o NPY está ausente de todos os tecidos năo apresentam alteraçőes demonstráveis no comportamento alimentar. O cruzamento reprodutivo entre camundongos com knockout de NPY e camundongos ob/ob revelou que a prole com duplo knockout tinha um fenótipo atenuado de obesidade. Estes dados indicam que o NPY é um regulador importante, mas năo exclusivo, do comportamento alimentar e gasto de energia.  FIGURA 15-11 FIGURA 15-11
Vários fatores periféricos que contribuem para o apetite podem agir sobre as açőes cerebrais [22]. Foi constatado que a grelina, que é sintetizada no estômago, desempenha um papel na iniciaçăo da refeiçăo, possivelmente através de açőes em neurônios-alvo no hipotálamo [23,24]. O peptídio YY é sintetizado no intestino e tem um efeito orexigęnico através da açăo no cérebro [25]. Agentes anoréticos também podem ser produzidos pelo intestino. O peptídio-1 semelhante ao glucagon (GLP-1) é sintetizado no intestino e age para suprimir o apetite em animais e seres humanos [26]. A colecistocinina (CCK), que é produzida nas células endócrinas, também age no sentido de inibir o apetite [27]. Finalmente, a insulina, que é produzida no pâncreas em resposta ŕs refeiçőes, pode também agir no cérebro para regular a ingestăo de alimentos e gasto de energia [28]. GR — receptor de grelina; LR — receptor de leptina. voltar
|
















;)
;)
;)
;)
;)
;)