  FIGURA 15-12 FIGURA 15-12
Causas monogęnicas de obesidade em roedores. A identificaçăo de causas monogęnicas de ocorręncia espontânea da obesidade ofereceu pistas significativas para compreender a regulaçăo do peso corpóreo. A identificaçăo do hormônio de gordura, a leptina [29], forneceu um mecanismo pelo qual a gordura pode sinalizar o estado dos depósitos periféricos de energia ao cérebro [30]. A leptina é também importante na regulaçăo de respostas fisiológicas ao jejum [31]. A análise de outros modelos de obesidade, como o camundongo Ay, forneceu um insight significativo das vias que săo importantes na regulaçăo do peso corpóreo. Modelos de engenharia tęm sido importantes na confirmaçăo da importância das vias e na identificaçăo dos papéis dos diferentes peptídios. AgRP — agouti-related peptide [peptídio relacionado ao agouti; CRH — hormônio liberador da corticotrofina; NPY — neuropeptídio Y.
 FIGURA 15-13 FIGURA 15-13
Causas monogęnicas da obesidade humana. Peptídios-chaves identificados em camundongos levaram ŕ busca de pacientes com defeitos semelhantes. Estes estudos foram feitos em pacientes com obesidade mórbida de início bem precoce. Em muitos casos (como deficięncia de leptina, deficięncia do receptor de leptina, anormalidades do receptor de melanocortina e mutaçőes de PC-1), o fenótipo humano é semelhante ao fenótipo de roedor. Algumas mutaçőes, porém, como as mutaçőes no gene POMC e as mutaçőes em PPARg2, só foram descritas em seres humanos.
 FIGURA 15-14 FIGURA 15-14
Causas monogęnicas de magreza em roedores. A maioria das causas monogęnicas da magreza de roedores foram lesőes especificamente criadas por engenharia. O mecanismo pelo qual algumas dessas manipulaçőes causam magreza é compreendido, como na superexpressăo hepática de leptina e deficięncia do hormônio concentrador de melanina [32]. Em outros casos, porém, como no knockout de proteína quinase A, năo se conhece o mecanismo de magreza. É digno de nota que a ablaçăo do gene NPY em roedores năo causou magreza nem quaisquer alteraçőes no fenótipo de alimentaçăo [33]. Entretanto, a ausęncia do neuropeptídio Y levou a uma atenuaçăo da obesidade encontrada no camundongo Lep ob /Lep ob [34]. As causas monogęnicas da magreza humana năo foram identificadas pois a busca de tais mutaçőes seria complicada. A definiçăo de critérios para a triagem de famílias apropriadas seria difícil porque os pesos, embora reduzidos, poderiam ainda estar dentro da faixa normal.
 FIGURA 15-15 FIGURA 15-15
Lista parcial de peptídios que regulam a ingestăo alimentar. Sabe-se que vários peptídios estimulam a ingestăo alimentar e que muitos mais a suprimem. Além disto, a ingestăo alimentar é regulada por monoaminas (năo discutidas aqui). CART — cocaine- and amphetamine-regulated transcript [transcrito regulado por cocaína e anfetamina].
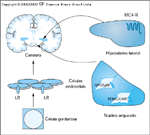 FIGURA 15-16 FIGURA 15-16
Sítios de lesőes de um único gene que levam ŕ obesidade humana. As relaçőes entre peptídios envolvidos na regulaçăo da alimentaçăo săo complexas. Esta figura sintetiza algumas das interaçőes conhecidas. A leptina é sintetizada por adipócitos e liberada na circulaçăo. O transporte de leptina através da barreira hematoencefálica é mediado pela forma curta do receptor da leptina (LR). No cérebro, a leptina tem como alvo os receptores de forma longa [35] tanto no hipotálamo arqueado como no dorsomediano (năo mostrado). No hipotálamo arqueado, a leptina regula os corpos celulares que co-expressam o peptídio relacionado ao agouti (AgRP) e o neuropeptídio Y (NPY) (peptídios que estimulam a alimentaçăo) e células que coexpressam a preproopiomelanocortina (POMC, o precursor do hormônio estimulante de melanócitos) e o transcrito regulado por cocaína e anfetamina (CART – cocaine- and amphetamine-regulated transcript) (peptídios que inibem a alimentaçăo). As células do arqueado projetam-se para várias áreas, inclusive o hipotálamo lateral [36], onde o receptor da melanocortina-4 (MC4-R) [37], que responde ŕ POMC e ao AgRP, é expresso. Este receptor pode estar presente em neurônios que expressam a orexina ou o hormônio concentrador da melanina, embora esta localizaçăo ainda năo tenha sido confirmada.
 FIGURA 15-17 FIGURA 15-17
A deficięncia de leptina causa a síndrome da obesidade encontrada nos camundongos ob/ob. Estes camundongos desenvolvem obesidade precoce associada ŕ hiperfagia e resistęncia ŕ insulina. A deficięncia de leptina foi descrita em duas crianças aparentadas. Ambas tiveram peso normal ao nascer, mas eram acentuadamente obesas durante a primeira infância. O eixo hipotálamo-hipófise-adrenal (HHA) era normal nestas crianças, embora o nível do hormônio estimulante da tireóide (TSH) estivesse ligeiramente elevada. Nas duas crianças, o controle glicęmico era normal; este achado difere da grave resistęncia ŕ insulina encontrada em camundongos. Os dois pares de pais eram de peso normal. A análise do gene da leptina revelou que os pais eram heterozigóticos e que as crianças eram homozigóticas para uma mutaçăo de deslocamento de quadro de leitura ('frameshift mutation') que levou ŕ síntese de espécies truncadas, năo-secretadas [38].
 FIGURA 15-18 FIGURA 15-18
Mutaçăo em receptor de leptina humano. A deficięncia de receptor da leptina foi descrita em uma família grande [39]. O probando (caso índice) apresentou-se com obesidade e hipogonadismo hipogonadotrófico significativos. Os homozigotos afetados tęm níveis de leptina aumentados 100 vezes (índice de massa corpórea [IMC]) a mais que o conteúdo gorduroso. Os heterozigotos afetados tinham IMCs na faixa pré-obesa ou minimamente obesa e níveis de leptina de aproximadamente 200 mg/mL.TSH - nível do hormônio estimulante da tireóide.  FIGURA 15-19 FIGURA 15-19
Peptídios envolvidos na regulaçăo do comportamento alimentar e pigmentaçăo. A intrigante conexăo entre pigmentaçăo e alimentaçăo foi sugerida pela primeira vez pelo achado que camundongos amarelos espontaneamente mutantes (Ay), conhecidos como camundongos agouti, também eram acentuadamente obesos. O agouti (normalmente expresso na pele) age sobre os melanócitos como um fator parácrino para inibir a conversăo de ficomelanina (amarelo) em eumelanina (preto). A maioria dos camundongos é castanha por causa de misturas variáveis destes dois pigmentos. Os camundongos Ay expressam agouti em todos os tecidos e de uma forma desregulada. Estes achados sugeriram que os receptores de melanocortina tęm um papel na mediaçăo do comportamento alimentar.
A proteína agouti é expressa na pele e regula a coloraçăo da pele, agindo através do receptor-1 da melacortina, onde inibe a açăo do hormônio estimulante de melanócitos; quando universalmente expressa, leva ŕ pigmentaçăo amarela (inibindo o receptor da melacortina-1) e ŕ obesidade por inibir os receptores de melanocortina-2 expressos centralmente. Estes receptores săo expressos no sistema nervoso central; quando ativados pelo hormônio estimulante de melanócitos, medeiam a inibiçăo do comportamento alimentar [40]. O hormônio concentrador da melanina regula a pigmentaçăo cutânea em peixes, é produzido na hipófise e liberado na circulaçăo e age sobre melanóforos (células contendo grânulos de pigmento) para causar agregaçăo de grânulos e escurecimento cutâneo. Năo tem nenhum papel conhecido na pigmentaçăo de mamíferos, mas, nos mamíferos, é produzido no hipotálamo lateral; estimula o comportamento alimentar através de um receptor ainda năo identificado. Um ligante antagonista endógeno potencial para os receptores de melanocortina hipotalâmica é a proteína relacionada ao agouti, um homólogo recentemente clonado do agouti. Esta proteína é expressa no núcleo arqueado do hipotálamo e seu RNA mensageiro é regulado para cima ('up-regulated') tanto nos camundongos ob/ob como nos db/db. A proteína relacionada ao agouti parece inibir o receptor de melanocortina de uma maneira semelhante ŕ do agouti.  FIGURA 15-20 FIGURA 15-20
O camundongo Ay, no qual a proteína agouti normal é expressa ectopicamente, é amarelo e obeso [41].
 FIGURA 15-21 FIGURA 15-21
Proteína agouti. A proteína agouti é normalmente expressa nos folículos pilosos da pele e regula a pigmentaçăo da pele e pelagem. A proteína agouti expressa perifericamente inibe o receptor da melanocortina-1 (MC1-R) e impede a conversăo do pigmento amarelo para o pigmento preto mediada pelo hormônio estimulante de melanócitos (MSH) [42]. Se for produzido na hipófise, o MSH regulará a pigmentaçăo; se produzido no hipotálamo, regulará a fome.
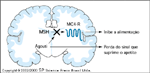 FIGURA 15-22 FIGURA 15-22
Proteína agouti. A proteína agouti (Ay) centralmente expressa age no receptor melanocortina-4 (MC4-R) [43] e impede a inibiçăo da alimentaçăo mediada pelo hormônio estimulante de melanócitos (MSH). A síndrome da obesidade poderia ser mimetizada pela obtençăo por engenharia genética de um camundongo no qual MC4-R estivesse ausente [44]; esta capacidade demonstra a importância deste receptor. No camundongo Ay, o agouti é expresso no cérebro. O agouti bloqueia o MC4-R, um dos dois receptores cerebrais de melanocortina. O MC4-R năo é capaz de responder ao a-MSH. Os efeitos inibitórios do a-MSH sobre a fome foram perdidos; portanto, o camundongo Ay é obeso.
 FIGURA 15-23 FIGURA 15-23
Expressăo do agouti e peptídio relacionado ao agouti (AgRP). Por năo ser o agouti normalmente expresso no sistema nervoso central, o achado do efeito do agouti sobre os receptores de melanocortina centralmente expressos levou a uma busca de peptídios semelhantes ao agouti no sistema nervoso central. O peptídio relacionado ao agouti [45,46] é expresso no núcleo arqueado e é um dos peptídios do sistema nervoso central que regulam o receptor da melacortina-4 (MC4-R). Além disto, as atençőes se concentraram sobre o hormônio estimulante de melanócitos-a (MSH), um produto da preproopiomelanocortina no arqueado, e sobre o papel do a-MSH na regulaçăo da alimentaçăo por agir como um agonista no MC4-R.
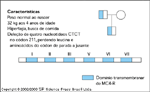 FIGURA 15-24 FIGURA 15-24
Mutaçőes no receptor da melacortina-4 (MC4-R). Uma coorte de crianças gravemente obesas foi submetida ŕ triagem de mutaçőes no MC4-R por meio de seqüenciamento direto de nucleotídeos [47]. Um paciente era heterozigótico para uma deleçăo de 4 pares de base no códon 211 do MC4-R. Esta mutaçăo resultou em um códon de parada ('stop codon') na regiăo que codifica o quinto domínio transmembranar. Os resíduos no quinto e sexto domínio transmembranar săo importantes para a sinalizaçăo do MC4-R e, portanto, esta mutaçăo resulta em um receptor năo-funcional. A măe do probando tem peso normal, mas o pai é obeso (índice de massa corpórea, 41). A mesma mutaçăo foi identificada no pai.
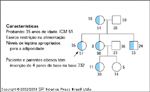 FIGURA 15-25 FIGURA 15-25
Mutaçőes de deslocamento de quadro de leitura ('frameshift mutations') do receptor da melanocortina-4 receptor (MC4-R) e obesidade humana herdada dominantemente. Uma populaçăo francesa foi submetida a triagem através da seleçăo de pessoas com uma história de obesidade na primeira infância e o maior índice de massa corpórea (IMC) de toda a vida, em qualquer dada idade [48]. O éxon único inteiro do MC4-R foi avaliado com o uso de cinco pares de primers. O seqüenciamento direto identificou um probando [caso índice] no qual uma mutaçăo de deslocamento de quadro de leitura heterozigótica resultou em um receptor truncado năo-funcional. A família do probando foi submetida ŕ triagem e foram identificados outros parentes com a mutaçăo. Todos esses parentes tinham níveis semelhantes de adiposidade. A idade dos indivíduos é indicada ao lado do símbolo e o IMC está abaixo do símbolo.
 FIGURA 15-26 FIGURA 15-26
Mutaçőes de POMC. As mutaçőes no gene POMC levam a uma síndrome de insuficięncia adrenal, pigmentaçăo vermelha de pęlos e obesidade. A preproopiomelanocortina (POMC) é o precursor de muitos peptídios, inclusive o hormônio adrenocorticotrófico (ACTH), hormônio estimulante de melanócitos (MSH) e b-endorfina. No paciente 1, duas mutaçőes distintas do POMC levaram ŕ interferęncia na síntese adequada de ACTH e MSH. A insuficięncia adrenal resulta da ausęncia de ACTH. O pęlo vermelho resulta da ausęncia da regulaçăo pelo MSH da pigmentaçăo no folículo piloso, que seria mediada pelo receptor da melanocortina-1. A obesidade resulta da ausęncia do MSH agindo centralmente, que regularia a alimentaçăo através do receptor da melanocortina-4 [41]. Uma síndrome de obesidade, insuficięncia adrenal e pigmentaçăo vermelha de pęlos foi encontrada em outro paciente descrito por Krude et al. [49]. Este paciente era homozigótico para uma mutaçăo que aboliu a traduçăo de POMC.
 FIGURA 15-27 FIGURA 15-27
Resumo das mutaçőes nas vias do sistema nervoso central que podem estar associadas ŕ obesidade em seres humanos. Setas azuis apontam para mutaçőes que foram identificadas. Setas cinzas com pontos de interrogaçăo apontam para os sítios em que podem ocorrer mutaçőes, mas que ainda năo foram confirmados. AgRP — peptídio liberador de agouti; BHE — barreira hematoencefálica; MC4-R — receptor da melacortina-4; MCH — hormônio concentrador da melanina; MSH — hormônio estimulante de melanócitos; PC1 — pró-hormônio convertase-1; POMC — preproopiomelanocortina; SSC — supressores da sinalizaçăo de citocina.
 FIGURA 15-28 FIGURA 15-28
Gene da pró-hormônio convertase 1 e obesidade. Foi descrito pelo menos um paciente gravemente obeso com uma mutaçăo na pró-hormônio convertase-1 (PC1). O paciente era um heterozigoto misto para o gene PC1 [50]. A síndrome clínica deste paciente era muito semelhante ŕquele encontrada no rato fa/fa. POMC — preproopiomelanocortina.
 FIGURA 15-29 FIGURA 15-29
PPAR-gama2. O receptor ativado pelo proliferador de peroxissomo (PPAR – peroxisome proliferator-activated receptor)-gama2 é um importante regulador da diferenciaçăo de adipócitos. Em um grande estudo de pacientes alemăes, foram identificados quatro pacientes obesos com uma mutaçăo "missense" no PPAR-gama2 [51]. Esta mutaçăo resultou na conversăo da prolina na posiçăo 115 para uma glutamina. A superexpressăo do gene mutante nos fibroblastos de camundongo revelou que a proteína mutante é defeituosa na fosforilaçăo de uma serina na posiçăo 114. Os fibroblastos que expressam o gene mutante exibiram uma diferenciaçăo acelerada em adipócitos. IMC — índice de massa corpórea.
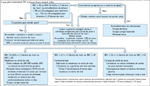 FIGURA 15-30 FIGURA 15-30
Algoritmo baseado em evidęncias para o tratamento da obesidade. IMC—índice de massa corpórea. (Adaptado de [52].)
voltar
|
















;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)