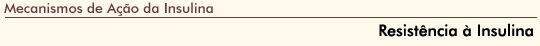
 FIGURA 2-15 FIGURA 2-15
Moduladores da açăo da insulina. Vários sinais convergem no receptor da insulina (IR – insulin receptor) para modular sua funçăo. As proteína-tirosina fosfatases (PTP – protein tyrosine phospha- tases) parecem desempenhar um papel importante na desativaçăo do IR, fornecendo um mecanismo para regular a resposta insulínica. A etapa endocítica que se segue ŕ ligaçăo com a insulina, com internalizaçăo do completo ligante/receptor para dentro das vesículas recobertas de clatrina, pode ser mais um ponto para a desfoforilaçăo do receptor. O endossomo foi identificado como um importante sítio de atividade da tirosina fosfatase com respeito ao IR [68]. Muitos estudos de seres humanos e roedores obesos resistentes ŕ insulina demonstram alteraçőes na expressăo e/ou atividade das PTPs nos tecidos responsivos ŕ insulina [69–73]. Há duas classes principais de tirosina fosfatases: de tipo receptor (ligada ŕ membrana) e de tipo năo-receptor (citosólica). As tirosinas fosfatases identificadas como IR fosfatases incluem LAR e LRP (de tipo receptor) bem como PTP1B (citosólicas). Estas fosfatases podem também alterar a sinalizaçăo da insulina definindo substratos IR como alvos de desfosforilaçăo. Estudos em camundongos transgęnicos revelam que uma superexpressăo de LAR em músculo resulta em resistęncia ŕ insulina no corpo todo [74]. Camundongos com knockout para PTP1B săo resistentes ŕ obesidade e exibem maior sensibilidade ŕ insulina [75,76]. Várias outras moléculas afetam a açăo da insulina. Foi proposto que a glicoproteína de membrana PC-1 exerceria um efeito negativo sobre a funçăo do IR por deprimir sua atividade de quinase [77]. Os estudos indicam uma correlaçăo entre a expressăo de PC-1 e a resistęncia ŕ insulina em paciente com tiabetes tipo 2 [78]. Outro inibidor da IR quinase é a glico-proteína sérica a2-HSG. Foi relatado que esta proteína de 63 kDa inibe a autofosforilaçăo do receptor induzida pela insulina, bem como a fosforilaçăo do substrato-1 do IR e do substrato do IR Shc, com uma reduçăo associada das açőes mitogęnicas da insulina. Năo houve relato de nenhum efeito da a2-HSG sobre as funçőes metabólicas da insulina [79]. Originalmente, a proteína semelhante ao ras associada ao diabetes (Rad – ras-like protein associated with diabetes) foi identificada por hidribizaçăo subtrativa como um gene superexpresso em pacientes com diabetes. Embora estudos subsqüentes năo tenham sido capazes de confirmar esta associaçăo, é interessante que a superexpressăo da Rad em células transfectadas comprometa a captaçăo de glicose insulino-dependente [80]. A glicoproteína transmembranar SHPS-1 é fosforilada e forma um complexo com a SHP-2, uma PTP năo-receptor, em resposta ŕ insulina. Este complexo parece intensificar a atividada da ERK em resposta ŕ insulina [81].
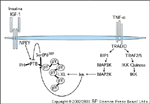 FIGURA 2-16 FIGURA 2-16
Fator-a de necrose tumoral (TNF-a) como modulador da açăo da insulina. A importante idéia de que a inflamaçăo está associada ŕ resistęncia ŕ insulina é conhecida há muito tempo [82] e é compatível com o achado de que citocinas induzidas por estresse, como TNF-a, causam resistęncia ŕ insulina. As cascatas de sinalizaçăo reguladas pelo TNF-a săo complexas e envolvem muitos pontos de ramificaçăo, inclusive a ativaçăo de várias serina quinases e fatores de transcriçăo que promovem a apoptose ou a proliferaçăo [83]. Uma das ramificaçőes da via de sinalizaçăo do TNF-a envolve a ativaçăo da c-Jun N-terminal quinase (Jnk) [84–86]. A Jnk é uma quinase induzida por estresse prototípica que é estimulada por muitos agonistas durante uma inflamaçăo aguda ou crônica. A Jnk fosforila inúmeras proteínas celulares, inclusive IRS-1 e IRS-2, Shc e Gab1 [87]. Um papel para a Jnk durante a açăo da insulina é forçoso, já que tanto o IRS-1 como o IRS-2 contęm motifs de ligaçăo ao Jnk. Este motif medeia a associaçăo específica da Jnk com o IRS-1, que promove a fosforilaçăo de um resíduo de serina específico que está localizado no lado COOH-terminal do domínio de ligaçăo ŕ fosfotirosina (PTB – phosphotyrosine-binding) (Ser307 no IRS-1 murino; Ser312 no IRS-1 humano). A fosforilaçăo deste resíduo inibe a funçăo do domínio PTB, que perturba a associaçăo entre o receptor da insulina e o IRS-1 e inibe a fosforilaçăo de tirosina [87]. Neste modelo, a ligaçăo do TNF-a com um receptor tipo 1 de TNF (TNFR1) resulta no recrutamento do fator 2/5 associado ao receptor de TNF (TRAF2/5 – TNF–receptor-associated factor 2/5) e proteína-1 de interaçăo com o receptor (RIP1 – receptor-interacting protein 1) através da proteína adaptadora TRADD. O TRAF2/5 e a RIP1 promovem a ativaçăo das proteína quinases Jnk e IkB quinase (IKK). A Jnk ativada se associa ao motif LXL de ligaçăo ŕ Jnk no IRS-1, que promove a fosforilaçăo de Ser307. A fosforilaçăo de Ser307 inibe a funçăo do domínio PTB e inibe a fosforilaçăo de tirosina estimulada pela insulina/IGF e a transduçăo de sinal.
voltar |
















;)
;)