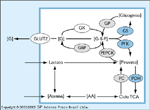
  FIGURA 8-6 FIGURA 8-6
Produçăo hepática de glicose. Esquema simplificado das principais vias intracelulares de produçăo de glicose no fígado, glicogenólise e neoglicogênese (linhas grossas). Formas ovais sombreadas indicam enzimas-chave sensíveis ŕ insulina na via. As concentraçőes de substratos estăo entre colchetes; as enzimas sensíveis ŕ insulina estăo dentro de círculos, sombreados para indicar açăo estimulante ou cinza para indicar açăo inibitória. Embora a degradaçăo de glicogęnio aumente diretamente as concentraçőes intra-celulares de glicose-6-fosfato (G-6-P), a captaçăo de lactato, alanina e outros aminoácidos neoglicogęnicos fornece os precursores de 3 carbonos para a síntese de novo de G-6-P. G — glicose livre; GP — glicogęnio fosforilase; GS - glicogęnio sintase; G6P — glicose-6-fosfatase; GK — glicoquinase; PFK — fosfofrutoquinase; PEPCK — fosfoenolpiruvato-carboxiquinase; PC — piruvato carboxilase; PDH — piruvato desidrogenase; AA - aminoácidos; GLUT2 — isoforma 2 (năo sensível ŕ insulina) do transportador de glicose;TCA — ciclo dos ácidos tricarboxílicos.
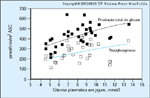 FIGURA 8-7 FIGURA 8-7
Contribuiçăo da neoglicogęnese para a concentraçăo plasmática de glicose em jejum. Produçăo endógena total de glicose (mensurada pela técnica da diluiçăo do rastreador) e neoglicogęnese (determinada pela técnica de água deuterada [6]) foram simultaneamente mensuradas em indivíduos năo-diabéticos em jejum e em pacientes com diabetes mellitus tipo 2. Como esperado (veja a Fig. 8-4), a produçăo endógena de glicose (quadrados cheios, linha sólida) está diretamente relacionada ao grau de hiperglicemia ao longo de uma faixa de concentraçőes plasmáticas de glicose em jejum. A neoglicogęnese (quadrados vazios, linha azul) gera aproximadamente metade da produçăo total de glicose em indivíduos năo-diabéticos e está aumentada em pacientes diabéticos, contribuindo, portanto, substancialmente para sua hiperglicemia de jejum [7]. ASC — área de superfície corpórea.
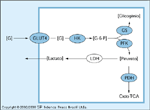 FIGURA 8-8 FIGURA 8-8
Captaçăo periférica de glicose. Esquema simplificado de síntese de glicogęnio e oxidaçăo de glicose, as principais vias da captaçăo intracelular de glicose nos tecidos-alvo da insulina. Formas ovais sombreadas indicam enzimas-chave sensíveis ŕ insulina na via. G — glicose livre; G-6-P — glicose-6-fosfato; GLUT4 — isoforma 4 (sensível ŕ insulina) do transportador de glicose; HK — hexoquinase II; GS — glicogęnio sintase; PFK — fosfofrutoquinase; PDH — piruvato desidrogenase; LDH — lático desidrogenase;TCA — ciclo dos ácidos tricarboxílicos.
 FIGURA 8-9 FIGURA 8-9
(Veja a Lâmina Colorida) Açăo da insulina no coraçăo. O músculo miocárdico é sensível ŕ insulina. Embora os ácidos graxos livres (FFA – free fatty acids) representem o combustível dominante para o músculo cardíaco no estado de jejum, um aumento nas concentraçőes circulantes de insulina inibe a lipólise, restringindo assim a disponibilidade de FFA e promovendo a captaçăo de glicose. Com o uso de 18F-desoxiglicose (FDG), um análogo da glicose (que é transportado e fosforilado da mesma maneira que a D-glicose, mas năo é adicionalmente metabolizado) marcado com um isótopo radioativo de vida curta do flúor (18F), a tomografia por emissăo de pósitrons (PET – positron-emitting tomography) detecta um sinal que é proporcional ŕ taxa de captaçăo miocárdica de glicose. A figura mostra imagens de FDG do músculo cardíaco humano durante um estudo de clamp euglicęmico de insulina, como aquele ilustrado na Figura 8-3. As cores (com vermelho sendo o mais intenso) indicam as regiőes com diferentes taxas de utilizaçăo da glicose. A, O ventrículo esquerdo de um paciente normal. B, As paredes ventriculares de um paciente que sofreu um infarto do miocárdio anterior 6 meses antes do estudo PET. Uma área "fria" de ausęncia de captaçăo de glicose está claramente visível no canto superior direito (a parede anterior do ventrículo esquerdo). Está também evidente uma diminuiçăo difusa na captaçăo da glicose mediada pela insulina (resistęncia ŕ insulina) por toda a parede ventricular esquerda, envolvendo regiőes miocárdicas distantes da área infartada e normalmente perfundida (mapeamento da perfusăo năo mostrado) [8].
 FIGURA 8-10 FIGURA 8-10
Mediçăo da captaçăo intracelular de glicose e do gasto de energia in vivo via calorimetria indireta. O esquema ilustra a correspondęncia entre calorimetria indireta (na qual consumo de oxigęnio = [O2 no ar expirado – O2 no ar inspirado] X fluxo de ar; produçăo de dióxido de carbono = [CO2 no ar expirado – CO2 no ar inspirado] X fluxo de ar) e o princípio de Fick (pelo qual o consumo de oxigęnio = (O2 em sangue arterial – O2 em sangue venoso central) X débito cardíaco; produçăo de dióxido de carbono = (CO2 em sangue arterial – CO2 em sangue venoso) X débito cardíaco). Com o uso de equaçőes calorimétricas, as taxas líquidas de oxidaçăo de lípides e carboidratos e de gasto de energia podem ser quantificadas no corpo inteiro, bem como no nível do órgăo começando com os dados de troca de gases. CE — coraçăo esquerdo; CD — coraçăo direito;VO2 — consumo de oxigęnio;VCO2 — produçăo de dióxido de carbono.
 FIGURA 8-11 FIGURA 8-11
Curva dose-resposta para açăo da insulina sobre a captaçăo oxidativa de glicose e năo-oxidativa de glicose no indivíduo saudável. A calorimetria indireta pode ser combinada com a técnica do clamp de insulina para estimar a oxidaçăo de glicose nos vários platôs de insulina plasmática. A captaçăo năo-oxidativa de glicose, consistindo primariamente na síntese de glicogęnio, é entăo obtida como a diferença entre a captaçăo total de glicose e a oxidaçăo líquida de glicose. A figura apresenta dados de indivíduos saudáveis estudados numa faixa de níveis plasmáticos de insulina; as linhas pontilhadas identificam os valores de Km para a captaçăo oxidativa de glicose e a síntese de glicogęnio. A oxidaçăo de glicose tem uma alta sensibilidade e uma baixa capacidade; a síntese de glicogęnio tem uma sensibilidade menor, mas uma capacidade maior. Portanto, sob condiçőes fisiológicas, uma hiperinsulinemia leve estimula a oxidaçăo de glicose, ao passo que uma insulinizaçăo mais forte promove o armazenamento de glicose no glicogęnio. Estudos de clamp combinados com calorimetria regional demonstraram que o músculo esquelético é o tecido-alvo da insulina responsável pela maior parte (50% a 70%) da captaçăo e armazenamento da glicose mediados pela insulina in vivo [9].
 FIGURA 8-12 FIGURA 8-12
Curvas dose-resposta para açăo da insulina sobre o metabolismo lipídico. A calorimetria indireta também fornece estimativas para a oxidaçăo líquida total de lípides. A oxidaçăo de ácido graxo livre (FFA – free fatty acid) circulante pode ser medida pela coleta de CO2 marcado no ar expirado durante a infusăo constante de palmitato marcado com carbono. A oxidaçăo lipídica tissular é entăo definida como a diferença entre a oxidaçăo lipídica total e a oxidaçăo de FFA. Nos estudos de clamp em indivíduos normais resumidos na figura, doses baixas de insulina inibiram efetivamente tanto a oxidaçăo lipídica total como a de FFA; doses fisiológicas altas de insulina (ou hiperinsulinemia crônica) năo afetaram adicionalmente a oxidaçăo de FFA, mas causaram síntese líquida de lípides (ie, valores negativos de oxidaçăo lipídica). PC — peso corpóreo. (Dados de Groop et al. [10].)
voltar |
















;)
;)
;)
;)
;)
;)
;)